Dinarokavkaski genom İ2/Eu7
Dinarokavkaski genom İ2/Eu7 (mutacija S31, prije: haplotip I1b1, I.Jurić: hrvatski haplotip), biokemijska formula: M170 - A>C327 tgcttcacacaaatgcgttt / ccaattactttcaacatttaagacc). Haplogrupa İ2/Eu7 (S31) je u ljudskoj genetici, haplogrupa muških Y-kromosoma u jugoistočnoj Europi i prednjoj Aziji. Njena veća i naša inačica tj. haplotip I2a2/Eu7 je tipična za naše dinarske i kavkaske etnogrupe, ponajviše kao 1/2 do 3/4 pučanstva za Hrvate, islamske Bošnjake, Crnogorce, kavkaske Swanete, Dargyne i kurdske Delamite (Zazaki), zatim manje oko 1/4 do 1/3 populacije za zapadne Srbe (jekavski Prečani), Moldavce, ine Kurde, Abhaze i Teheran, a rjedje s nižim udjelom za ostale susjedne narode jugoistočne Europe i prednje Azije tj. Starog istoka. Druga manja inačica ili İ2a1/Eu8 (ranije: I1b2), je u visokom udjelu specifična za otočne starosjedioce na Sardiniji i s manjim udjelima u Baskiji, Italiji, Francuskoj, itd. Nasuprot učestalosti na jugoistoku Europe i u prednjoj Aziji podgrupe I2 i na jugozapadu inačice I2b, varijanta I1 se može najčešće naći kod pučanstva sjeverozapadne Europe, osobito u Švedskoj, Norveškoj, Danskoj, Njemačkoj, Nizozemskoj itd.
Sadržaj
Abstract
Dinaric-Caucasian haplogroup İ2/Eu7: In human genetics, Haplogroup İ2 is a Y-chromosome haplogroup. Until 2008, it was known as Haplogroup I1b. Haplogroup I2 might have originated in Southeastern Europe some 15,000 - 17,000 years ago and developed into five main subgroups : İ2*, I2a1, I2a2, I2b1 and I2b2. Haplogroup I2a1 (M26) accounts for approximately 40% of all patrilines among the Sardinians. It is also found at low to moderate frequency among populations of the Pyrenees (Baskians), and it has been found in 9.4% of a sample of Albanians living in FYR Macedonia. The age of YSTR variation for the M26 subclade is 8.0±4.0 kya. I2a2I2a2a (P41.2) is typical of the West Balkans populations in south-eastern Europe, being highest in Croatia and Bosnia-Herzegovina. Haplogroup I2a2a is also found in north-eastern Italians. There is also a high concentratio of I2a2a in Moldova, north-east Romania and western Ukraine. The subclade divergence for P37.2 occurred 10.7±4.8 kya. The age of YSTR variation for the P37.2 subclade is 8.0±4.0 kya. The age of YSTR variation for the M423 subclade is 8.8±3.6 kya. Pericic places its expansion to have occurred "not earlier than the YD to Holocene transition and not later than the early Neolithic”. However, according to Ken Nordtvedt I2a2a arose 2500 years ago in Eastern Europe. In 2010 Nordtvedt suppoused that I2a2a is just too young to not have been a result of a sudden expansion. Nordtvedt has suppoused that around 15,000 years ago the two branch-lines of I2a2 (M423) eventually leading to I2a2a (P41.2) and I2a2b (L161) separated.
Nadgrupa İ/Eu7-8 i podgrupe
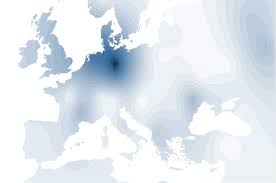
Veliki dio tj. oko 1/4 današnjih Europljana pripadaju široj nadgrupi I/Eu7, koja je jedna je od najvećih haplogrupa u srednjoj i jugoistočnoj Europi te oko Kavkaza. Ova je najčešća i dominantna kod Hrvata pa je bila nazvana hrvatskom ili dinarskom, ali je bila pronađena u visokom postotku i kod Skandinavaca i Nijemaca, a mnogo Sardinaca nosi njenu varijantu s dodatnom mutacijom nazvanu I2b/Eu8. Nositelji I/Eu7 su najvjerojatnije doselili u jugoistočnu Europu iz jugozapadne Azije prije ledenog doba, što znači pred više od 20.000 godina i odatle se proširili po srednoj, sjevernoj i istočnoj Europi. Daljim istraživanjima se otkrilo kako je I/Eu7 razgranata mutacijama u 4 glavna i više manjih podogranaka. Glavne pripadne haplogrupe su: izvorna I*, I1a, I2a i I2b. I1a i c su najbrojnije u izvorno germanskim zemljama a manje u krajevima koje su naselila germanska plemena: Normandija, Engleska i na širem okružju Moskve – možda su ju tamo donesli srednjovjeki Varjagi (Vikinzi) iz Skandinavije. I2a i c su česte kod Kozaka oko ušća Dnjestra.
Korijenska podgrupa I* je nađena raspršeno po cijeloj Europi i prednjoj Aziji s niskim udjelom, osim što je malo više ima u Turskoj i Francuskoj (oko 5%). I1a je značajno prisutna u srednjoj i sjeverozapadnoj Europi i Skandinaviji. Na istom području je najveća koncentracija I1c, ali ona čini manji postotak nego I1a. Naša haplogrupa I2a je najčešća u jugoisočnoj Europi, a ima je dosta još u srednjoj i istočnoj Europi. Među pripadnim narodima je ima jednako ili više od I*, I1a i I1c zajedno. Oko Kavkaza skoro svi Y kromosomi haplogrupe I spadaju u I2a podgrupi. Iz I2 je mutirala podgrupa I2b/Eu8 kojoj pripada većina muškog pučanstva Sardinije odakle se manje proširila na susjedne obale i dalje u zapadnu Europu. Jedino među Baskima I2b se nalazi u malo većem postotku. Haplogrupa I/Eu7 je dijelom karakteristična za Europu i izvan nje je prisutna u većem postotku među Kurdima i na Kavkazu, pa u Turskoj i Iranu. Zanimljivo je da su u Turskoj nađene sve glavne podgrupe uključivo izvornu I*. I1c je nađen sporadično u Pakistanu.
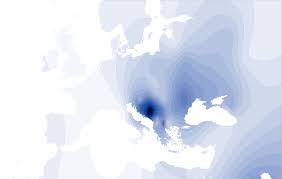
Nadgrupa İ/Eu7-8 obuhvaća europske i prednjoazijske populacije s najvišim udjelom od Skandinavije preko srednje Europe i Balkana do Kavkaza i Teherana. Kako ta nadgrupa većinom obuhvaća nepodobne narode iz europske Osovine u 2. svj. ratu i još u Aziji iz "Osovine zla", to je odnedavna njena genetika izložena raznim kontroverznim postupcima na ime 'političke korektnosti' i inih neznanstvenih interesa. Pripadne populacije se danas ugrubo biokemijski dijele u 2 geografske haplogrupe: sjeverno-germanska İ1 u Skandinaviji i srednjoj Europi, ter naša dinarokavkaska İ2 u jugoistočnoj Europi i prednjoj Aziji. Iako je njihovo razdvajanje najvjerojatnije nastalo u postglacialu, bez sigurnih pokazatelja o tomu se istraživači pobliže ne slažu o vremenu razdvajanja, za koje razni auktori predlažu izmedju 12.000 do 3.000 god. starosti. Naša haplogrupa İ2/Eu7 (P37.2) se pretežno nalazi kod pučanstva jugoistočne Europe i Kavkaza, s najvećom učestalošću osobito u Dalmaciji i u Hercegovini (50 - 73 %). Veliki udjel ove haplogrupe na dinarskom području se možda može objasniti time što je presušeni manji Jadran pri oledbi poslužio kao toplije sklonište za pučanstvo ove podgrupe.
Nama bliska srodna podgrupa İ2b/Eu8 (P41.2 / M359) je najviše nadjena kao dominantna na otoku Sardiniji i još s puno manjim udjelom kod Baska. Više od 40% Sardinaca pripada ovoj grupi. Izumrlo zapadnoeuropsko paleolitsko pučanstvo koje je nosilo haplogrupu İ2b (M26) je moralo preživiti oledbu negdje zapadno od Apenina u južnoj Francuskoj ili zapadnoj Italiji, odakle je pred oko 9.000 godina uspješno naselilo Sardiniju u doba najniže morske razine. Unatoč tomu što je haplogrupa I2b (P41.2=M359) dominantna na Sardiniji, a potječe od dominantno dinarske haplogrupe I2a-P37.2, podgrupe I2b nema istočno od Francuske i Italije iako je s manjim udjelom nađena još u pučanstvu Baleara, Kastilije, Baskije, Pirineja, jugozapadne Francuske, itd. Stoga je ta zapadna podgrupa I2b u bliskoj vezi s jugozapadnim Europljanima paleolitskog iskona i njihovi nosioci imaju tek dalju vezu s dinarskim pučanstvom haplogrupe I2a.
Dinarokavkaska haplogrupa İ2/Eu7 (biokemijska formula: M170 - A>C327 tgcttcacacaaatgcgttt / ccaattactttcaacatttaagacc) se nalazi uglavnom na Balkanu kao i na Kavkazu, a najviše u Hrvatskoj 45% do 66% i u BiH 48 – 73% gdje ima najveću poznatu koncentaciju u Hercegovini (Marjanović i sur. 2005). Kod nas ga ima najviše u Hercegovini, Dalmaciji i istočnoj Slavoniji, a najmanje u Kvarneru i Zagorju, pa tom genomu uglavnom pripada dinarski tip pučanstva BiH, te srednje i južne Hrvatske. Znakovito je kako se Eu7 (M-170) pojavljuje u disjunkciji tj. dalje na istoku izdvojeno u Moldaviji (32 %) i još izvan Europe najviše oko Kavkaza (34-82%), pa u kaspijskom Dagestanu i središnjoj Aziji gdje niz tamošnjih etnogrupa sadrže po 10 - 25% Eu7, što je možda u vezi s dalekim praiskonom ranih pradinaraca (neolitski "Hurrduw").
Ovo je zamalo jedini europski genom čiji posebni jezični ekvivalent dosad još nije jasno odredjen (kod nas je to vjerojatno starodalmatski jezik), ali njihov izvorni jezik sigurno nije slavenski kojemu pripada drugi genetski tip R1a/Eu19. Danas toj haplogrupi u Europi uglavnom pripadaju razne inačice naših ikavaca: ikavski šćakavci, ikavski kajkavci i jadranski čakavci, a izrazito najviši udjel i apsolutnu većinu taj tip dostiže na otocima, npr. na otoku Hvaru gdje ima većinu od 2/3 (66 %), pa 55% na Braču, 54% na Korčuli, 45% na dalmatinskom kopnu i najviše u Hercegovini 73%. Na slično takodjer upućuje poredba grobnih kostura u Dalmaciji i Hercegovini iz antike i srednjeg vijeka koji ne pokazuju slavenske promjene nego uglavnom nastavak dinarskih tipova (Mikić 1976, 1979, 1984), te arheonalazi Komanske kulture u Dalmaciji i Crnoj Gori iz 6.- 9.st. od Zadra do Skadra, koji još ne pripadaju Slavenima nego starosjediocima (Vinski 1971, Marčinko 2000).
Etnički udjeli İ2/Eu7
Dinarokavkaska haplogrupa İ2/Eu7 (M 170) je manjeviše obilno zastupljena u jugoistočnoj Europi i prednjoj Aziji od Teherana do Slovenije (usp. Semino & al. 2000, Wells & al. 2001, Underhill & al. 2001, Rootsi & al. 2004, Nasidze & al. 2003, 2004, 2005, Jurić 2003-2011):
- Azijski genski dinarci: najviše kavkaski Swaneti 82% i Dargyni 58%, Abhazi 34%, Teheran 33%, Delamiti (Zazaki) 33%, Oseti 32%, Gagauzi 31%, Andi 27%, Adygejci 25%, Mordvini 19%, Isfahan 17%, Tatari 16%, Taškent 12%, Čuvaši 11 %, Buhara 10%, ini Kurdi 9%, Pakistan 8%, Uzbeki 7%, Udmurti 7%, Karačaji 7%, Nenezi 6%, Fergana 5%, Azeri 5%, Lezgini 5%, Nogajci 5%, Sirijci 5%, Libanon 5%, Armenci 4%, Kazbeki 4%, Marijci 4%, pravi Turci (bez Kurda) 4% - (ini Azijati su ispod 4%) i kod američkih Indianaca 2%.
- Europski genski dinarci: Hercegovina 73%, Dalmacija 56%, sva Hrvatska (prosjek) 46%, slavonski Šokci 42%, Bosna 42%, Crna Gora 38%, Slovenija 38%, Rudanovi "Jugohrvati" (bez Like i Dalmacije) 34%, Moldavija 32%, Magjarska 28%, Albanija 24%, Kozaci 23%, Rumunji 22%, Makedonci 20%, otok Kreta 18%, Poljaci 18%, Bukovina 18%, Ukrajinci 18%, Andaluzija (jug Španjolske) 14%, sva Grčka (prosjek) 14%, 'Čehoslovaci' 14%, otok Sicilija 12%, Rusi (srednji) 11%, Magjari 11%, Švicarska 8%, Italija (kopno) 7% - (ini Europljani su ispod 7%).
Hrvatski genski dinarci
Naš dominantni dinarski haplotip İ2/Eu7 (tj. po novom sinonimu I-M170) kod Hrvata je nazočan prosječno sa 37 - 49%, a nadprosječno iznad 1/2 pučanstva (50-73%) osobito u Hercegovini 73%, Dalmaciji 50-68%, Lici, Kordunu, Banovini i donjoj Posavini (Šokci) 42%. Znakovito je da se osim prednje Azije i najviše Kavkaza (vidi niže), ovaj još manje nalazi (2 %) i kod američkih Indianaca, što upućuje na moguće pradavne kontakte dinaraca i Indianaca. Izrazito najbrojnija haplogrupa Y-kromosoma u Hrvatskoj po svim dosad provedenim studijama je I2. Njen udjel u pučanstvu je od 40-73% tj. 2/5 do 3/4 populacije, ovisno o području i studiji. U drugoj studiji (Peričić et al, 2005) su analizirani Y-kromosomi iz BiH, Srbije, Makedonije i Kosova, ali uz idejno-podobni dio Hrvatske bez neslavenske Like i Dalmacije. Osim među Hrvatima, haplogrupa I2 ima nadpolovičnu većinu među bosanskim Y-kromosomima, oko 1/3 svih Y-kromosoma Srba i Makedonaca i 20% Slovenaca. Kao i u inim studijama, najbrojnija je podgrupa I2a iako je u svim populacijama nazočna i podgrupa I1 tipična za sjevernu u zapadnu Europu. Samo među Hrvatima su nađene sve 4 glavne podgrupe iz I. Zanimljivo je da su na Krku i Braču nazočne uz najveću I2, još i ostale podhaplogrupe u visokom postotku za naše krajeve: 18% na Krku i 17% na Braču. Nažalost taktički nije navedeno o kojim podgrupama se radi. I2a ima takodjer visoki udjel 42% kao dominantna i na sjeveru medju slavonskim Šokcima (I. Jurić 2005, 2007, 2011).
Azijski dinarci İ2/Eu7
Iz gornjeg popisa je očevidno kako naš dinarski haplotip İ2/Eu7 uopće nije ograničen samo na Europu, kako to uporno prikrivaju, zaobilaze i prešućuju zamalo svi naši i zapadnjački autori iz idejno-pristranih poriva. Zbog oskudnih istraživanja u Aziji prije 2003. nije bio sigurno potvrdjen izvan Europe, ali po vjerodostojnim analizama Rusa i inih (iz bivšeg SSSRa), on se manjeviše nalazi iznad 5% kod dvadesetak raznih naroda diljem jugozapadne i srednje Azije, a kod nekih oko Kavkaza je podjednako obilan kao kod nas pa nikako nije ograničen u Europi kako neki žele. Ipak već Semino i sur. (2000) i A. Gibbon (2000) ispravno naslućuju da je pripadna naša mutacija M-170 ovamo stigla ranijim selidbama negdje iz “Srednjeg Istoka” (jugozapadne Azije), što su nakon 2001. dosad jasno potvrdili niže navedeni biogenetski nalazi osobito iz Kavkaza (do 58% Eu7), Teherana i Kurdistana (33-34% Eu7), a s manjim udjelom i dalje na istoku sve do Fergane i Pakistana: usp. Weale i sur. 2001, Wells i sur. 2001, Quintana-Murci i sur. 2002, Nasidze i sur. 2004. i 2005, itd.
Kavkaski dinarci
Dinaridne etnogrupe Kavkaza: Dargyni, Swaneti, Abhazi, Osseti, Adygeji, Karačaji itd. (usp. Ačkasova i sur. 1990, Zerjal i sur. 2002, Nasidze i sur. 2003, 2004, 2005, itd.). Glavni i najizrazitiji biogenetski centar s jasnom i za Aziju najvećom nazočnošću našega dinarskog haplotipa je veliki masiv Kavkaza, gdje ga sadrži cijeli niz desetak raznih reliktnih naroda s arhajskim jezicima. Izrazito najviši udjel našega dinarskog haplotipa tu imaju kavkaski Swaneti čak 82%, Dargyni 58%, Abhazi 33%, Osseti 32%, Adygeji 25% Eu7, pa još Kubani 16%, Karačaji 8%, Lezgini 5% i većina inih susjednih naroda na Kavkazu ispod 5%. Tu je na zapadnokavkaskom području Kubana takodjer nadjen i karirani hrvatski grb na keramičkom skifosu iz II. stoljeća (Ačkasova i sur. 1990). Ove dinaroidne genske etnogrupe zapadnog Kavkaza danas žive na jugoistoku Azovskog mora, tek par desetaka kilometara južnije od antičkog Tanaisa gdje su nadjene 2 antičke ploče s grčkim natpisima našeg etnonima Horouathos (Latyšev 1890).
Dinaridni Kurdi
Delamiti i Kurmandji (Kandler 1984, Paul 1995, Zerjal i sur. 2001, Al-Zahery i sur. 2003, Cinnioglu i sur. 2004, Nasidze i sur. 2005, Issa-Fatimi i Yoshamya 2009): Donedavna je genetika Kurda bila skoro nepoznata, ali nam nove analize daju vrlo znakovite pokazatelje za iskon Hrvata. Kod južnih Kurda u Iraku izrazito dominiraju semitski haplotipovi (Eu9 i Eu10), dok je tu naš I2/Eu7 daleko rjedji pa njegov prosjek za Irak iznosi tek 0,7%. Naprotiv spram sjevera njegov udjel medju Kurdima naglo raste, pa je u istočnoj Anatoliji kod najveće kurdske grupe Kurmandji koja obuhvaća blizu polovice svih Kurda, njegov prosjek već iznosi 16%. Dapače, medju sjevernim kurdskim plemenima (Dimili, Zazaki, Kizilbaši) naš I2/Eu7 doseže još veći udjel do 33% tj. skoro kao i kod nas u Hrvatskoj. Ovi sjeverni Kurdi su izvan Europe u Aziji i jezično razmjerno najbliži Hrvatima, čak sa 28% sličnih izoglosa u njihovom rječniku, još uz nazočnost niza naših fonetskih i gramatičkih osobitosti: glasovi “jat”, č, ć, ž itd. (Issa-Fatimi i Yoshamya 2009).
Takodjer i povijesna vrela (Menitsky 1964, Paul 1995) pokazuju nam da ovi sjeverni Kurdi potječu od klasične etnogrupe Delamiti iz pokrajine Dailam na sjeverozapadu Irana i tu je znakovita sličnost s antičkim Delmatima, na čijem je području u Hercegovini i danas najviši europski udjel našeg haplotipa Eu7 čak sa 73%. Inače su Kurdi jedan od najstarijih povijesno dokumentiranih naroda prednje Azije gdje žive još od halkolitske predhistorije, pa ih u prvim klinopisima navode već najraniji Sumerani kao sjeverniju zemlju Karda čak prije 5 milenija, pa Asirci pred 4 tisućljeća kao susjedna gorska plemena Kurki i napokon stari Grci kao planinski narod Kardouchoi (Xenophont god. 394.-371. pr. Kr.). Stoga naš dinarski haplotip u Kurdistanu najvjerojatnije postoji barem kroz 5 tisućljeća pa su odonda jamačno isključene njegove doselidbe iz Europe (što ideološki žele neki iliristi). Zato je povijesno i biogenetski puno vjerojatnija baš obratna selidba iz Kurdistana spram Dalmacije i Hercegovine, koja je bar dijelom povezana s osvajanjima staroperzijskih Ahemenida na Balkanu u 6.st. pr. Kr. (Ćurić 1991), nakon čega i grčki pomorci po prvi puta navode pleme Delmata u Dalmaciji, gdje su oni tada potisnuli druga starija ilirska plemena.
Iranski dinarci
Teheran i Iran (usp. Nasidze i sur. 2003, 2004, Qamar i sur. 2002, Quintana-Murci i sur. 2001): Slično Kurdima se naš I2/Eu7 nalazi najviše na sjeverozapadu i sjeveru Irana, a sve manje južnije i na jugoistoku. Tako je npr. u devetmilijunskoj prijestolnici Teheranu nadjena čak 1/3 ili 34% tog tipa, a južnije u Isfahanu 17%. Stoga brojčano u samom Teheranu danas živi više genskih dinaraca negoli kod nas u cijeloj Hrvatskoj, što je paralelno s povijesnim pokazateljima kulturnih sličnosti.
Dinarski Kozaci
Donski Kozaci (usp. Rootsi i sur. 2004): Na jugoistočnoj granici Europe i Azije, uz donji tijek rijeke Don i oko antičke luke Tanais (kod današnjeg Rostova) žive donski Kozaci kao potomci donedavnih konjaničkih nomada. Takodjer i u njihovom genomu je ondje nadjen naš dinarski haplotip sa 17% I2/Eu7. Dakle je danas Tanais na sjeveru i jugu okružen populacijama u kojima je manjeviše vidljivo nazočan naš dinarski haplotip s udjelom od 17 – 58%. Obzirom da se taj haplotip i sada nalazi uz bivši Tanais i još obilnije južnije uzduž Kavkaza, jedva može biti sumnje da je isti bio manjeviše zastupljen i medju antičkim Sindo-Meotima na istoku Azovskog mora, pa jamačno i kod naše rane etnogrupe Horouathos u samom Tanaisu s kojime tako direktno imamo odnedavna i biogenetsku, a ne samo imensku vezu.
Srednjoazijski dinarci
Taškent, Buhara i Fergana (Wells i sur. 2001, Zerjal i sur. 2002): Još istočnije od najobilnijeg Kavkaza i Kurdistana, na širjem prostoru stepa i polupustinja u Turkestanu do središnje Azije, tj. u Turkmeniji, Kirgiziji i Tadjikistanu je ta naša dinarska haplogrupa takodjer manjeviše difuzno rasprostranjena većinom u nižim udjelima ispod 15%, što je vjerojatno posljedica prastarih susjedskih primjesa s prahrvatskim pretcima. Npr. u Turkmeniji je za Taškent poznato 12% i za Buharu 10%, a kod još desetak naroda srednje Azije je njegov manji udjel od 4%-5%. Najistočnije dosad poznato nalazište naše dinarokavkaske haplogrupe u središnjoj Aziji je dolina Fergana u gorju Tyan-Shan sa 5% I2/Eu7. Stoga je taj dinarokavkaski haplotip manjeviše raširen na prostranom području šumostepa i polupustinja prednje i srednje Azije, od Kavkaza i Kurdistana sve do Fergane, što upućuje na približna područja ranijih selidbi naših predaka ili bar nama najbližih biogenomskih srodnika.
Indovedski dinarci
Krajnje najjužnije područje Eurazije, gdje je dinarokavkaska grupa I/Eu7 dosad bolje poznata u značajnom udjelu je sjeverozapadni dio Pakistana (Tribal Area) u graničnom pojasu uz Afganistan (klasična Harauvatya), gdje je kod više plemena nadjeno 7%-16% Eu7 (Qamar i surad. 2002).
Podvale s İ2/Eu7 i R1a/Eu19
Kad su prve detaljne analize Y-haplogrupa Hrvata od god. 2000-2004. (D. Primorac, M. Marcikić, P. Rudan, I. Jurić i dr.) konačno otkrile kako medju svim jezičnim Slavenima imaju baš Hrvati nedopustivo najmanje (23-27%) 'slavenskog' haplotipa (Dalmacija tek 9-20%) - da se povrh toga još usudimo imati čak osobiti dominantni haplotip İ2/Eu7 prosječno 37-46% (Dalmacija i Hercegovina čak 60-73%), usljedio je vrlo fanatični protuudar jugoslavista i jugonostalgika, uz logističko-financijsku podršku vanjskih imperialnih mentora koji su podržavali propalu Jugoslaviju i moguću obnovu nove 'Balkanije' i 'Jugosfere'. Prvi imperialni plotun protiv te 'politički nekorektne' genetike nepodobnih Hrvata je ispalio časopis 'Nature' pamfletom s potpisom mikrobiologa dr. M. Radmana kojim se traži prekid i korekcija tih istraživanja (dakako u jugo-smjeru). Zatim su provjerenom metodom (mrkve + batine) obećane znatne potpore za protivna 'istraživanja' i sinekure da se to poništi, što je prihvatio Antropološki institut u Zagrebu i od 2005. su svojim objavama poništili vlastite ranije rezultate kao i inih genetičara.
Taktika te naručene javne podvale je ova: iz tzv. 'hrvatskog' genskog prosjeka su izbačeni i izostavljeni idejno-nepodobni nalazi Like i Dalmacije s premalo slavenskog R1a i previše I2 (tim gorje po činjenice), pa sad politički korektno 'glajhšaltani' novi jugo-Hrvati već imaju podnošljivih 33% R1a + 34% I2: Taj je brainwash od 2005. nadalje javno i uzastopno obznanjen, pa citiran kroz niz časopisa i ponavljan popularnom filmskom emisijom kroz TV. Time Hrvati postaju genetski podobni i prikladni za Jugosferu i Balkaniju, pa Antropološki institut dobiva poveća sredstva i opremu, a organizator prof. P.Rudan postaje akademikom. Doduše, protiv toga javno protestiraju drugi, kao i sveučilišni genetičar prof.dr. Ivan Jurić koji je novim analizama potvrdio i proširio ranije rezultate do 2004. i prokazao krivotvorbe,- ali bez učinka jer "psi laju-karavana prolazi" tj. očite krivotvorbe se uporno nastavljaju u podobno-naručenu geostratešku pseudoznanost. Nakon te uspješne podvale kod nas, slično se s nepodobnom grupom I/Eu7 odnedavna ponavlja i u prednjoj Aziji, gdje je odjednom u novijim američkim analizama "netragom nestala" iz Kavkaza, Kurdistana i Irana,- gdje je već prije obilno nadjena u ruskim, njemačkim i inim izvješćima.
Antropološki odraz İ2/Eu7
Narodi s ovom dominantnom nadgrupom (I/Eu7) danas govore raznim jezičnim skupinama, a u Europi pretežno germanski. Morfološki se ističu izrazito vitkim i najvišim rastom od svih Europljana - nadasve Dinarci (I2) i Skandinavci (I1). Kosa im je ravna, pretežno brinet-kestenjasta i oči sivozelene do smedjaste, a brada uža i izbočena. Osobiti oblik razmjerno glomazne lubanje je često specifičan uglato-izdužen poput kutije zaobljenih uglova (tzv. "ćoškaste glave"), dok im je snažan "orlovski" nos razmjerno najveći medju Europljanima. Dinarski starci većinom imaju specifičnu dvodjelnu ćelavost u obliku potkove (tzv. "fratarska tonzura"), gdje rano ćelavljenje počinje bočno iznad oba oka i kružno oko glave, dok u sredini na tjemenu uglavnom preostaje dlakavi otok. Medicinski se nosioci İ2 medju Europljanima najviše ističu kao dugoživi stoljetni metuzalemi (Kavkaz, Sardinija itd.), ali su od inih osjetljiviji na neke bolesti npr. na HIV pa brže stradaju od AIDS-a nego ostali.
Iliri i provale Slavena
Od 6. stoljeća započinju prodori Slavena preko rimskih provincija i u tom naletu propadaju mnogi gradovi, imanja, utvrde i hramovi. Ugledni rimski historičar Procopius u zapisima iz 550. godine je opisao Slavene kao nevidjene sirove divljake i najveću opasnost po dotadanju civilizaciju, navodeći precizne podatke o svim njihovim pokoljima Ilira i Tračana. Slavenske pljačkaške horde predvodjene Avarima su bile toliko nemilosrdne da je staro domaće pučanstvo bilo prinuđeno prihvatati njihov jezik kako bi bar prividnom asimilacijom privolili avaro-slavenske bande da poštede živote pučanstva. Izrijekom se navodi da je u tim pokoljima pobijeno ili odvedeno u roblje preko 4 milijuna ljudi ili većina tadanjeg pučanstva rimske provincije Dalmacije tj. Ilirije. Od tih čestih slavenskih pokolja autohtonog pučanstva opisan je npr. masakr preko 7.000 žena i djece za samo dva dana 548. godine u gradu Draču (Dyrachion) što navodi Procopius. Naši ideologizirani jugo-istoričari redovito taktički zaobilaze, prešućuju i izostavljaju Prokopija i ine tadanje rimske izvore, pa od 6.- 8. stoljeća lažno tvrde kako im tada navodno "nedostaju istorijski izvori", samo da bi onda uljepšali i uzveličali tobože slavnu i miroljubivu "Veliku Seobu Slavena".
Medjutim,- znanost ipak ide dalje i bar modernim neutralnim i egzaktnim metodama forenzike se jasno dokazuju zamalo sve prozirne obmane, koje su nam silom 'mozgopranja' desetljećima i stoljećima nametnuli raniji okupatori uključivo prije Slavene i novije Jugoslavene. Taj ključni dokaz protiv jugoslavističkih idejnih podvala je danas prvenstveno biokemijska genomika, uz ine suvremene metode koje uglavnom istosmjerno opovrgavaju povijesne podvale zadojenih jugoslavista i sada ponovo materialno-forenzički dokazuju zamalo sve navode Prokopija i onodobnika o koljačkim provalama Slavena (bez stvarne selidbe), - Ove su bile bar podjednake ili još daleko gorje od kasnijih turskih, četničkih i sličnih novijih napada što su doista čisti naivci u poredbi s "romantičnim" Starim Slavenima. Iz toga danas proizlaze 3 ključna rezultata koji nas neizbježno sile na radikalnu reviziju i odbacivanje u ideološku ropotarnicu dosad izmišljene i neistinite "pseudoistorije jugoslavista":
- 1. Kao prvo, u zadnjih desetak godina je više nego jasno i uzastopce potvrdjeno po Y-haplotipu, kako u najvećem dijelu Hrvatske i BiH, a ponajviše u Hercegovini, Liki i Dalmaciji ima čudnovato malo Slavenima sličnog haplotipa Eu19/R1a: u cijeloj Hrvatskoj i Bosni slavenski biogenski tip obuhvaća tek 1/4 do 1/3 pučanstva tj. od 27-35% genskih Slavena medju nama, a samo rubno na sjeverozapadu npr. u Vojvodini, Podravini, Zagorju, Istri, Gorskom Kotaru i Sloveniji je nešto više do polovice genskih Slavena (Semino i sur. 2000, I.Jurić 2003, 2005, 2007, 2011). Dapače, u Hercegovini, Liki i Dalmaciji je udjel neslavensko-dinarskog haplotipa Eu7/I1b još viši oko 1/2 do 3/4 ili od 46-72% pučanstva, a ostalo su razni ini tipovi pa je tu slavenski genski udjel zanemarivo neznatan - ali isti genski neslaveni tu sad govore slavenski. Ovo znači isto što piše Prokopije: bili su nasilno "poslavenjeni". To dalje još znači da u tim dinarskim krajevima zapravo nije bilo tzv. "Velike Seobe Slavena".
- 2. Kao drugo i najvažnije je poredbeni omjer muških i ženskih slavenskih gena kod nas, što nam sada dubinski otkriva čak i tip same doselidbe. Kod normalne selidbe naroda, plemena i obitelji je završni omjer muško-ženskih gena bar približno podjednak. Takva se genska ravnoteža kod nas donekle i vidi - ali samo u spomenutim rubnim sjeverozapadnim krajevima i najviše u Sloveniji i Makedoniji gdje su slavenski haplotipovi muškog Y-tipa i ženskih slavenskih mitohondrija podjednako zastupljeni: Ovo znači kako je barem te rubne krajeve izvan Dinarida zahvatila normalna pučka "Selidba Slavena" tj. miroljubivih agrarnih doseljenika. Naprotiv, u većem dijelu Hrvatske je prosječno preko 90% ženskih mitohondrija kod nas neslavenskog iskona, a jedva 6% - 9% ženskih Hrvatica su iskonom Slavenke, što konačno znači da je u Hrvatskoj 3 do 4 puta manje genskih Slavenki od manjinskih muških Slavena. Dapače baš u istim Dinarskim krajevima Like, Hercegovine i Dalmacije slavenske Hrvatice genski umalo ni ne postoje. To stvarno znači upravo ono što piše Prokopije i suvremenici: da su uglavnom muški Slaveni u rimsku Dalmaciju ili Ilirik doista stizali kao razbojničke koljačke horde (bez svojih žena i obitelji), pa su najvjerojatnije tu stvarno pljačkali uz genocidne pokolje i bar tu u Iliriku uglavnom nije ni bilo "Velike Seobe Slavena" s 'miroljubivim' seljacima, kako nas već desetljećima lažno obmanjuju jugoistoričari i naši slavisti.
- 3. Treće i ključno je, da u našemu središnjem povijesnom etnoprostoru tj. u Dalmaciji i BiH gdje uglavnom nema slavenskih mitohondrija, naprotiv postoji obilje specifičnih azijatskih mitohondrija koji su najčešći u jugozapadnoj Aziji tj. u bivšemu Perzijskom carstvu. Oni su kod tih središnjih Hrvatica češći negoli igdje u Europi, osim još kod Bugarki i Laponaca. To zapravo znači da u srednjovjeki hrvatski etnoprostor nije doselila samo neka manja vojna elita Iranohrvata, nego cijeli jedan kompletni Iranohrvatski narod sa ženama i djecom. Dakle: krajem antike i u ranomu srednjem vijeku je na dinarski prostor i istočni Jadran genetski stigla velika doselidba Iranohrvata, a ne navodna idejno napuhana "Seoba Slavena" od koje su sada preostale samo pljačkaške provale muških Slavena bez žena i naroda.
Inačice i sinonimi
Simboličke medjunarodne oznake za tu haplogrupu (i većinu inih) još nisu ujednačene ni standardne, pa se svakih par godina zajednički mijenjaju. Dosad postoje najmanje 3 uzastopna sustava imenovanja, dok pojedini auktori i istražni timovi imaju još i svoje dodatne oznake.
İ2 (L68, M438/P215/S31) (prije: I1b)
- İ2a (P37.2) (prije: I1b1) = prvotno Eu8 - Sardinija i Baski
- I2a*
- I2a1 (M26) (prije: I2a2, I1b1b)
- I2a1*
- I2a1a (M161) (prije: I2a2a, I1b1b1)
- I2a2 (M423)(prije: I2a1)
- I2a2a (P41.2/M359.2) (prije: I2a1a, I1b1a)
- I2a2b (L69)
- İ2b (M436/P214/S33, P216/S30, P217/S23, P218/S32) (prije: I1b2) = prvotno Eu7 - Balkan itd.
- İ2b*
- I2b1 (M223, P219/S24, P220/S119, P221/S120, P222/U250/S118, P223/S117) (prije: I1b2a, old I1c)
- I2b1*
- I2b1a (M284) (prije: I1b2a1)
- I2b1a1(L126/S165, L137/S166)
- I2b1b (M379) (prije: I1b2a2)
- I2b1c (P78) (prije: I1b2a3)
- I2b1d (P95) (prije: I1b2a4)
- I2b2 (L38/S154, L39/S155, L40/S156, L65/S159)
Literatura
- Ivan Jurić 2003: Genetičko podrijetlo Hrvata. Vlastita naklada, Zagreb (2. izdanje: Slobodna Dalmacija, Split 2005).
- Ivan Jurić 2005: On the Y Chromosome Haplotype of the First Farmers in the Historical Territory of Croatia and the Directions of Agricultural Diffusion in Europe, Agric. conspec. sci. 70/4: 121-126.
- Ivan Jurić 2007: Genetičko podrijetlo šokačkih rodova na području Vinkovaca. Godišnjak ogranka Matice Hrvatske sv. 24, Vinkovci.
- Ivan Jurić 2011: Genetika Hrvata. Zagreb (u tisku).
- Ivan Nasidze, Schadlich H., Stoneking M. 2003: Haplotypes from the Caucasus, Turkey and Iran for nine Y-str loci. Forensic Sci. Int. 137: 85-93.
- Ivan Nasidze & al. (17 koautora), 2004: Mitochondrial DNA and Y-Chromosome variation in the Caucasus. Ann. Human Genet. 68: 205-221.
- Ivan Nasidze & al. 2005: MtDNA and Y-Chromosome variation in Kurdish groups. Annals of Human Genetics, 69: 401-412.
- Mirabal S., Varljen T. & al. 2010: Human Y-chromosome short tandem repeats, A tale of acculturation and migrations as mechanisms for the diffusion of agriculture in the Balkan Peninsula. Amer. Journal of Physic. Anthrop. 142/3: 380–390. doi:10.1002/ajpa.21235. PMID 20091845
- Kr. Rebala, A.I. Mikulich & al. (7 koautora) 2007: Y-STR variation among Slavs, evidence for the Slavic homeland in the middle Dniepr basin. Journal Human Genet. Japan.
- Rootsi S. & al. 2004: Phylogeography of Y-chromosome haplogroup I reveals distinct domains of prehistoric gene flow in Europe. Am. J. Hum. Genet. 75/1: 128–37. doi:10.1086/422196. PMC 1181996. PMID 15162323
- Barać L, Pericić M, Klarić IM, et al. (July 2003). "Y chromosomal heritage of Croatian population and its island isolates". Eur. J. Hum. Genet. 11 (7): 535–42. doi:10.1038/sj.ejhg.5200992. PMID 12825075
- Rootsi Siiri & al. 2004: Phylogeography of Y-chromosome Haplogroup I reveals Distinct Domains of Prehistoric Gene Flow in Europe. Amer. Journal of Human Genet. 75/1: 128–137. doi:10.1086/422196. PMC 1181996. PMID 15162323. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1181996
- D. Marjanovic & al. 2005: The peopling of modern Bosnia-Herzegovina (Y-chromosome haplogroups in the three main ethnic groups). Ann. Hum. Genet. 69/6: 757–763. PMID 16266413, doi:10.1111/j.1529-8817.2005.00190.x.
- Alexander Varzari 2006: Population History of the Dniester-Carpathians, Evidence from Alu Insertion and Y-Chromosome Polymorphisms. Dissertation, Facul. Biol. Ludwig-Maximilians University, München.
- Tatiana Karafet & al. 2008: New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree. Genome Research, doi:10.1101/gr.7172008.
- Vincenza Battaglia & al. 2008: Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe. European Journal of Human Genetics; doi: 10.1038/ejhg.2008.249.
- Ken Nordtvedt: "The Lichtenstein cave Ydna haplotypes show three from the new S23+(xM223) I2b* (ISOGG 2008) tree".
Vanjske sveze
- Phylogeography of Y-Chromosome Haplogroup I (Rootsi 2004)
- Rescalled Haplogroup I Tree (K. Nordtvedt 2011).
- Frequency and Variance of I2 (called I1b)
- Y-DNA Haplogroup I and Its Subclades from the International Society of Genetic Genealogy (ISOGG)of 2008
- Phylogeography of Y-Chromosome Haplogroup I
- Frequency Distributions of Y-DNA Haplogroup I and its subclades - with Video Tutorial
- Frequency and Variance of I1b (now considered I2a2-M26)
- Map of 'I1a' (now considered I1-M253)
- Map of 'I1b' (now considered I2a-P37.2)
- Map of 'I1c' (now considered I2b-M223)
{kind=link}
{kind=link}
{kind=link}
Poveznice
- Genomi neslavenskih Hrvata
- Mitohondrijska DNK Hrvatica
- Eurozapadni genom R1b/Eu18
- Eurazijski genom R1a/Eu19
- Orwelska pseudo-genetika
- Biogenomika Perzije
Reference
Condensed compilation from above literature and links, adapted and elaborated by GNU-license almost from WikiSlavia and Wikinfo.